ECOLINK-Salix (Sweden-Germany)
The three experimental field sites of ECOLINK-Salix have been established along a 1500 km latitudinal gradient, from Uppsala (Sweden) to Rostock and Freiburg (Germany), in spring 2014. In 2020 the Freiburg site was abandoned. Now the ECOLINK-Salix experiment consists of two sites. The plantations are short rotation coppice stands. Shoot biomass was harvested for the first time after three years of growth (in winter 2016/2017). The overall goal of ECOLINK-Salix is assessing the effects of genotype identity and diversity in willow short rotation coppice on various ecosystem functions and, ultimately, tree growth and yield stability (e.g. resistance to drought and pests). Particular focus lies on functions that are important for ecological sustainability, such as water and nutrient use, carbon sequestration, and biodiversity.
 |
| The location of the site in Uppsala, Rostock, and Freiburg |
Design
The experiment has a randomized block design, with three replicates (blocks). Salix varieties, i.e., genotypes, with known characteristics and different taxonomic relationships (full siblings to different species) were planted in all theoretically possible combinations: plots with single varieties, mixtures of two, three, and four varieties. The diversity treatments were assigned randomly to plots within a block. Four Salix varieties were planted - from near to more remote taxonomic relationship:
- 'Tora' (Svalöf-Weibull (SW) cultivar no. 910007, S. schwerinii × S. viminalis)
- 'Björn' (SW 910006, Salix schwerinii E. Wolf. × S. viminalis L.)
- 'Jorr' (SW 880013, S. viminalis)
- 'Loden' (SW 890129, S. dasyclados)
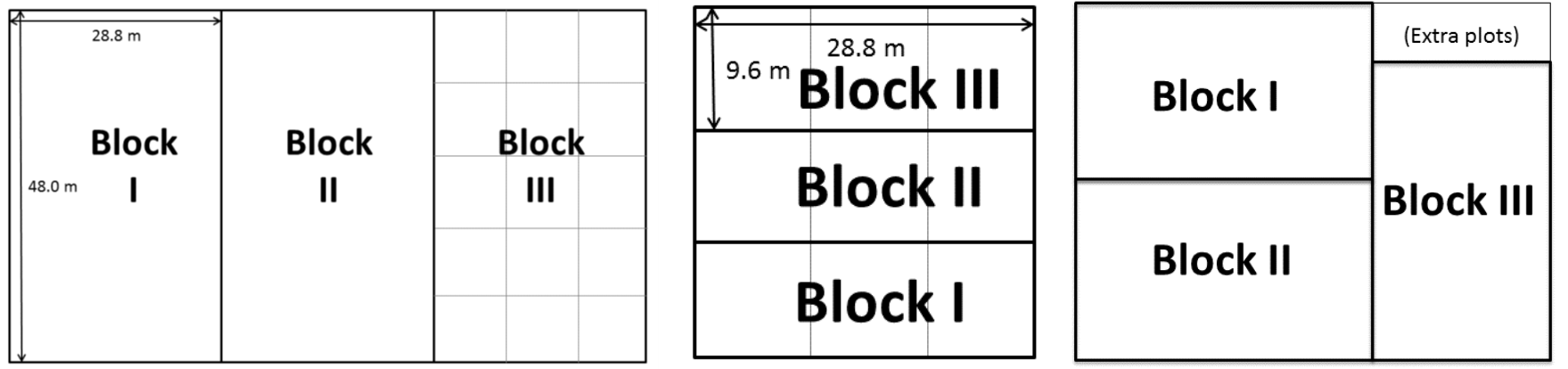 |
| The blocks at Uppsala, Rostock, and Freiburg. A block in Uppsala or Freiburg contains 15 plots (1-4 varieties); a block in Rostock contains 3 plots (1-2 varieties). All plots are 9.6 m x 9.6 m large. |
 |
| Hexagonal planting design in plots with one, two, and three varieties. In plots with three and four varieties, the position of varieties was randomized, but within columns and lines, each individual is directly neighboured by an individual of a different variety. Planting density was c. 15600 plants ha-1. Each plot contains nine subplots (3.2 m x 3.2 m, 16 plants), which will be used for specific assessments, e.g., litter decomposition, weed dynamics, soil biology. |
Site characteristics
| site | Uppsala | Rostock | Freiburg (abandoned in 2020) |
|---|---|---|---|
| former land use | arable | arable | grassland (sheep grazing) |
| altitude | 12 m | 13 m | 230 m |
| soil type | vertic cambisol | stagnic cambisol | cambisol |
| area | 4147 m2 | 829 m2 | 4424 m2 |
| no of plots | 45 | 9 | 45 |
| plot size | 92.16 m2 | 92.16 m2 | 92.16 m2 |
| no of trees planted | 6480 | 1296 | 6480 |
| planting date | 2014 | 2014 | 2014 |
| diversity variables | genetic diversity | genetic diversity | genetic diversity |
| diversity gradient | 1-4 varieties | 1-2 varieties | 1-4 varieties |
| size species pool | 4 | 2 | 4 |
| species pool | Salix varieties: Tora, Björn, Jorr, Loden |
Salix varieties: Tora, Loden |
Salix varieties: Tora, Björn, Jorr, Loden |
| contact person | Martin Weih | Christel Baum | Michael Scherer-Lorenzen |
Research
We will test the ecosystem consequences of adding individual genotypes of trees. We want to functionally and quantitatively explore how the heritable traits of some genotypes of Salix affect (1) ecosystem productivity, litter decomposition and soil carbon sequestration; (2) the abundance and structure of associated mycorrhiza communities; (3) the action of herbivorous insects; and (4) the diversity and function of understory vegetation. At individual sites, additional research questions will be conducted, e.g. on resource use complementarity, soil hydrology or wood quality. The research will have implications for future bio-energy concepts because new genotypes of energy crops such as Salix generated by plant breeding must comply with sustainability requirements, e.g., biodiversity and carbon sequestration, for a carbon-neutral and environmentally friendly energy source, e.g. Weih et al. (2014).
Watch this talk for a summary of our research on species or genotype identiy vs. stand diversity in Salix biomass plantations for productivity and other ecosystem functions (26th IPC session, October 2021).
Extra information
Send an e-mail to the contact persons or explore the publications:
theses
- Arranz C (2020) Use of mixtures in short rotation coppice plantations to increase productivity. PhD thesis, University of Freiburg
- Hoeber S (2019) Biodiversity-ecosystem function in a willow biomass production system. PhD thesis, Uppsala: Sveriges lantbruksuniv., Acta Universitatis Agriculturae Sueciae, 1652-6880 (link)
- Lindh M, Hoeber S, Weih M, Manzoni S (2022) Interactions of nutrient and water availability control growth and diversity effects in a Salix two-species mixture. Ecohydrology e2401 https://onlinelibrary.wiley.com/doi/10.1002/eco.2401
- Hoeber S, Baum C, Weih M, Manzoni S, Fransson P (2021) Site-dependent relationships between fungal community composition, plant genotypic diversity and environmental drivers in a Salix biomass system. Frontiers in Fungal Biology, section Fungi-Plant Interactions 2: 671270 - https://doi.org/10.3389/ffunb.2021.671270
- Koczorski P, Furtado BU, Gołȩbiewski M, Hulisz P, Baum C, Weih M, Hrynkiewicz K (2021) The effects of host plant genotype and environmental conditions on fungal community composition and phosphorus solubilization in willow short rotation coppice. Frontiers in Plant Science 12: 647709 - https://doi.org/10.3389/fpls.2021.647709
- Koczorski P, Furtado B, Hrynkiewicz K, Breezmann M, Weih M, Baum C (2021) Site-effects dominate the plant-availability of nutrients under Salix species during the first cutting cycle. Forests 12: 1226 - https://doi.org/10.3390/f12091226
- Kollberg I, Weih M, Glynn C (2021) The effect of willow diversity on insect herbivory and predation. Agricultural and Forest Entomology (online) - https://doi.org/10.1111/afe.12466
- Kwon T, Shibata H, Kepfer-Rojas S, Schmidt IK, Larsen KS, Beier C, Berg B, Verheyen K, Lamarque J-F, Hagedorn F, Eisenhauer N, Djukic I, TeaCompositionNetwork (2021) Effects of climate and atmospheric nitrogen deposition on early to mid-term stage litter decomposition across biomes. Frontiers in Forests and Global Change 4: 678480 - https://doi.org/10.3389/ffgc.2021.678480
- Messier C, Bauhus J, Sousa-Silva R, Auge H, Baeten L, Barsoum N, Bruelheide H, Caldwell B, Cavender-Bares J, Dhiedt E, Eisenhauer N, Ganade G, Gravel D, Guillemot J, Hall JS, Hector A, Hérault B, Jactel H, Koricheva J, Kreft H, Mereu S, Muys B, Nock CA, Paquette A, Parker JD, Perring MP, Ponette Q, Potvin C, Reich PB, Scherer-Lorenzen M, Schnabel F, Verheyen K, Weih M, Wollni M, Zemp DC (2021) For the sake of resilience and multifunctionality, let's diversify planted forests! Conservation Letters e12829 - https://doi.org/10.1111/conl.12829
- Weih M, Nordh N-E, Manzoni S, Hoeber S (2021) Functional traits of individual varieties as determinants of growth and nitrogen use patterns in mixed stands of willow (Salix spp.). Forest Ecology and Management 479 - https://doi.org/10.1016/j.foreco.2020.118605
- Hoeber S, Fransson P, Weih M, Manzoni S (2020) Leaf litter quality coupled to Salix variety drives litter decomposition more than stand diversity or climate. Plant and Soil 453: 313-328 - 10.1007/s11104-020-04606-0
- Schweier J, Arranz C, Nock C A, Jaeger D, Scherer-Lorenzen M (2019) Impact of Increased genotype or species diversity in short rotation coppice on biomass production and wood characteristics. BioEnergy Research 12: 497-508 - https://doi.org/10.1007/s12155-019-09997-2
- Weih M, Glynn C, Baum C (2019) Willow short-rotation coppice as model system for exploring ecological theory on biodiversity–ecosystem function. Diversity 11(8): 125 - https://doi.org/10.3390/d11080125
- Hoeber S, Arranz C, Nordh N-E, Baum C, Low M, Nock C, Scherer-Lorenzen M, & Weih M (2018) Genotype identity has a more important influence than genotype diversity on shoot biomass productivity in willow short-rotation coppices. GCB Bioenergy, 10(8): 534-547 - https://doi.org/10.1111/gcbb.12521
- Djukic I, Kepfer-Rojas S, Kappel Schmidt I, Steenberg Larsen K, Beier C, Berg B, Verheyen K, TeaComposition (2018) Early stage litter decomposition across biomes. Science of the Total Environment
- Baum C, Hrynkiewicz K, Szymanska S, Vitow N, Hoeber S, Fransson PMA, Weih M (2018) Mixture of Salix genotypes promotes root colonization with dark septate endophytes and changes P cycling in the mycorrhizosphere. Frontiers in Microbiology 9: 1012 - doi: 10.3389/fmicb.2018.01012
- Müller M, Klein A-M, Scherer-Lorenzen M, Nock CA, Staab M (2018) Tree genetic diversity increases arthropod diversity in willow short rotation coppice. Biomass and Bioenergy 108: 338-344 - doi:10.1016/j.biombioe.2017.12.001
- Hoeber S, Fransson P, Prieto-Ruiz I, Manzoni S, Weih M (2017) Two Salix genotypes differ in productivity and nitrogen economy when grown in monoculture and mixture. Frontiers in Plant Science 8: 231 - doi: 10.3389/fpls.2017.00231
 |
 |
 |
| The Uppsala site during the third growing season (May 2016), the Rostock site at the time of establishment (April 2014), and the Freiburg site after the third growing season (October 2016) | ||
Funding
The ECOLINK-Salix project was funded by the Swedish Energy Agency (project no. 36654-11 and 36654-2) and the Swedish Research Council Formas (projects no. 2016-00998 and 2020-02339). In addition, the trial in Freiburg was supported financially through the "Innovationsfonds Forschung" of the Freiburg University for the project "Mechanisms of tree diversity effects on ecosystem functioning, and through the "Forschungsprogramm Bioökonomie" of the state of Baden-Württemberg for the project "Hochdivers-KUP: Nutzung von Baum-Diversität zur Erhöhung der Produktivität und ökologischen Stabilität von Kurzumtriebsplantagen".