The Sardinilla biodiversity experiment is one of the oldest experiments in TreeDivNet. The goal is to study complex links between biodiversity, land use and tropical ecosystem functioning, with a focus on biogeochemistry and plant health. Sardinilla consists of two subexperiments: the 'main' plantation (established in 2001) and the 'high diversity' one (established in 2003). The Sardinilla experiment was ended in the summer of 2018.

|
|
one of the plots in the main plantation in 2002
|
Design
main plantation
The 6 native tree species planted are of local economic and ecological value. They were chosen based on their relative growth rates: from fast-growing to more slowly growing long-lived pioneer species. The plantation consists of 12 monocultures (2 for all 6 species), 6 plots of different 3-species mixtures (1 species of the 3 growth rate categories), and 6 replicates of the 6-species mixture. Diversity treatments were randomly distributed to plots to reduce bias caused by differences in soil conditions.
Within a plot, planting followed a multiple Latin-square design to ensure that systematic environmental variation did not bias the results. The seedlings were planted in diagonal rows, with 3 m planting distance. The species identity of neighbours around trees of a given species thus remained the same within a plot. The planting density was similar to standard reforestation practices in Central America, which allows comparing with other plantations
high-diversity plantation
The 8 plots follow a split-plot design with 3 diversity levels (6, 9 and 18 species) per plot. The highest species number is comparable to the tree species diversity in intact natural forests (19 species) and was chosen based on the estimated tree species diversity of 45 m x 45 m plots in the nearby lowland tropical moist forest of Barro Colorado Island. The diversity levels are nested within 4 different species composition lines, which makes it possible to test the effect of species composition, accumulation, and diversity on ecosystem functioning. The regular design within the plots also allows for an explicit test of neighbourhood effects.
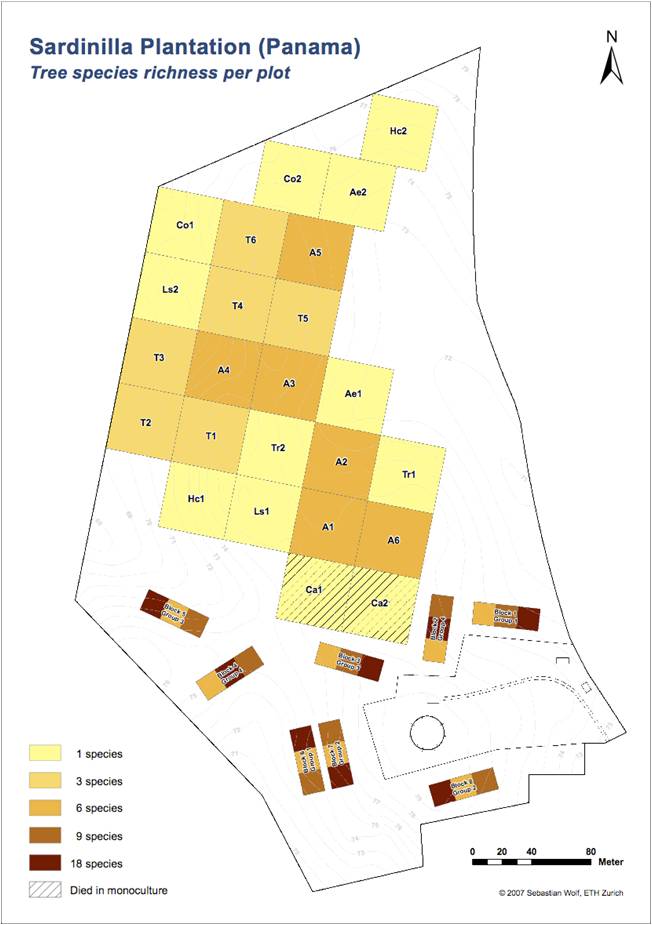 |
The 24 plots of the main plantation (1, 3, 6 species)
and the 8 plots of the high-diversity plantation (6, 9, 18 species) |
Site characteristics
| subexperiment |
main plantation |
high-diversity plantation |
| location |
Sardinilla |
| former land use |
original forest logged in 1952-53, 2 years of agriculture, then pasture |
| altitude |
70 m |
| soil type |
clay Alfisol, bedrock of limestone and other sediments |
| area |
9 ha |
| no of plots |
24 |
8 |
| plot size |
45 m x 45 m |
15 m x 45 m |
| no of trees planted |
5566 |
864 |
| planting date |
July 2001 |
July 2003 |
| diversity variables |
species richness
identity functional group |
species richness
species assemblage identity |
| diversity gradient |
1, 3, 6 sp.
3 growth rates |
6, 9, 18 sp.
4 assemblages |
| size species pool |
6 |
26 |
| species pool |
fast-growing: Cordia alliodora, Luehea seemannii
intermediate: Anacardium excelsum, Hura crepitans
slowly-growing: Cedrela odorata, Tabebuia rosea |
Albizia adinocephala, Anacardium excelsum, Astronium graveolens, Bursera simaruba, Cordia alliodora, Calycophyllum candidissimum, Colubrina glandulosa, Cedrela odorata, Dipteryx panamensis, Dalbergia retusa, Diphysa robinioides, Enterolobium cyclocarpum, Erythrina fusca, Gliricidia sepium, Guazuma ulmifolia, Hura crepitans, Hymenaea courbaril, Inga punctate, Luehea seemannii, Ormosia macrocalyx, Pachira quinata, Pseudomanea saman, Spondias mombin, Terminalia Amazonia, Tabebuia rosea, Vochysia ferruginea |
| contact person |
Catherine Potvin |
Research
The experiment was specifically designed to test how increases in tree species richness affect the cycling and storage of carbon. The research goal is to study the complex links between biodiversity, land use and tropical ecosystem functioning, with a focus on biogeochemistry and plant health. The high-diversity plantation was specifically planted to answer the following questions: (1) Is the 6-species diversity treatment relevant to the natural level of tree diversity in the local forests? (2) What is the importance of community composition at high tree species diversity?
Studies focus on the influence of tree species diversity on plant productivity, biomass accumulation, carbon sequestration potential, and carbon cycling; biogeochemistry, e.g., nitrogen and phosphorus dynamics, and water cycling; element cycles and water use of individual trees; plant health and herbivory. As tree-to-tree interactions are highly important to understand diversity effects, the impact of neighbour trees is studied in detail.
Extra information
Send an e-mail to the contact person, visit the experiment's own website, or explore the publications:
- Cesarz S, Craven D, Auge H, Bruelheide H, Castagneyrol B, Gutknecht J, Hector A, Jactel H, Koricheva J, Messier C, Muys B, Brien MJO, Paquette A, Ponette Q, Potvin C, Reich PB, Scherer-Lorenzen M, Smith AR, Verheyen K, Eisenhauer N (2022) Tree diversity effects on soil microbial biomass and respiration are context dependent across forest diversity experiments. Global Ecology and Biogeography 31:872-885 - https://doi.org/10.1111/geb.13461
- Kunert N, Brändle J, El-Madany TS (2022) Carbon allocation and tree diversity: shifts in autotrophic respiration in tree mixtures compared to monocultures. Biologia - https://doi.org/10.3389/fevo.2022.808589
- Duarte MM, Moral RdA, Guillemot J, Zuim CIF, Potvin C, Bonat WH, Stape JL, Brancalion PHS (2021) High tree diversity enhances light interception in tropical forests. Journal of Ecology 109(7): 2597-2611 - https://doi.org/10.1111/1365-2745.13669
- Madsen C, Kunz M, von Oheimb G, Hall JS, Sinacore K, Turner BL, Potvin C (2021) Influence of neighbourhoods on the extent and compactness of tropical tree crowns and root systems. Trees 35: 1673-1686 - https://doi.org/10.1007/s00468-021-02146-3
- Messier C, Bauhus J, Sousa-Silva R, Auge H, Baeten L, Barsoum N, Bruelheide H, Caldwell B, Cavender-Bares J, Dhiedt E, Eisenhauer N, Ganade G, Gravel D, Guillemot J, Hall JS, Hector A, Hérault B, Jactel H, Koricheva J, Kreft H, Mereu S, Muys B, Nock CA, Paquette A, Parker JD, Perring MP, Ponette Q, Potvin C, Reich PB, Scherer-Lorenzen M, Schnabel F, Verheyen K, Weih M, Wollni M, Zemp DC (2021) For the sake of resilience and multifunctionality, let's diversify planted forests! Conservation Letters e12829 - https://doi.org/10.1111/conl.12829
- Proß T, Bruelheide H, Potvin C, Sporbert M, Trogisch S, Haider S (2021) Drivers of within-tree leaf trait variation in a tropical planted forest varying in tree species richness. Basic and Applied Ecology 55: 6-19 - https://doi.org/10.1016/j.baae.2021.01.009
- Guillemot J, Kunz M, Schnabel F, Fichtner A, Madsen CP, Gebauer T, Härdtle W, von Oheimb G, Potvin C (2020). Neighbourhood‐mediated shifts in tree biomass allocation drive overyielding in tropical species mixtures. New Phytologist - https://doi.org/10.1111/nph.16722
- Madsen C, Potvin C, Hall J, Sinacore K, Turner BL, Schnabel F (2020) Coarse root architecture: Neighbourhood and abiotic environmental effects on five tropical tree species growing in mixtures and monocultures. Forest Ecology and Management 460:117851 - https://doi.org/10.1016/j.foreco.2019.117851
- Schnabel F, Schwarz JA, Dǎnescu A, Fichtner A, Nock CA, Bauhus J, Potvin C (2019) Drivers of productivity and its temporal stability in a tropical tree diversity experiment. Global Change Biology 25: 4257-4272 - https://doi.org/10.1111/gcb.14792
- Grossman JJ, Vanhellemont M, Barsoum N, Bauhus J, Bruelheide H, Castagneyrol B, Cavender-Bares J, Eisenhauer N, Ferlian O, Gravel D, Hector A, Jactel H, Kreft H, Mereu S, Messier C, Muys B, Nock C, Paquette A, Parker J, Perring MP, Ponette Q, Reich PB, Schuldt A, Staab M, Weih M, Zemp DC, Scherer-Lorenzen M, Verheyen K (2018) Synthesis and future research directions linking tree diversity to growth, survival, and damage in a global network of tree diversity experiments. Environmental and Experimental Botany 152: 68-89 - doi: 10.1016/j.envexpbot.2017.12.015
- Guerrero-Ramírez NR, Craven D, Reich PB, Ewel JJ, Isbell F, Koricheva J, Parrotta JA, Auge H, Erickson HE, Forrester DI, Hector A, Joshi J, Montagnini F, Palmborg C, Piotto D, Potvin C, Roscher C, van Ruijven J, Tilman D, Wilsey B, Eisenhauer N (2017). Diversity-dependent temporal divergence of ecosystem functioning in experimental ecosystems. Nature Ecology & Evolution 1(11): 1639-1642 - doi: 10.1038/s41559-017-0325-1
- Guerrero-Ramírez NR, Craven D, Messier C, Potvin C, Turner BL, Handa IT (2016) Root quality and decomposition environment, but not tree species richness, drive root decomposition in tropical forests. Plant and Soil 404: 125-139 – https://doi.org/10.1007/s11104-016-2828-y
- Verheyen K, Vanhellemont M, Auge H, Baeten L, Baraloto C, Barsoum N, Bilodeau-Gauthier S, Bruelheide H, Castagneyrol B, Godbold D, Haase J, Hector A, Jactel H, Koricheva J, Loreau M, Mereu S, Messier C, Muys B, Nolet P, Paquette A, Parker J, Perring M, Ponette Q, Potvin C, Reich P, Smith A, Weih M, Scherer-Lorenzen M (2016) Contributions of a global network of tree diversity experiments to sustainable forest plantations. Ambio 45: 29-41 - doi: 10.1007/s13280-015-0685-1
- Paine CET, Amissah L, Auge H, Baraloto C, Baruffol M, Bourland N, Bruelheide H, Daïnou K, de Gouvenain RC, Doucet J-L, Doust S, Fine PVA, Fortunel C, Haase J, Holl KD, Jactel H, Li X, Kitajima K, Koricheva J, Martínez-Garza C, Messier C, Paquette A, Philipson C, Piotto D, Poorter L, Posada JM, Potvin C, Rainio K, Russo SE, Ruiz-Jaen M, Scherer-Lorenzen M, Webb CO, Wright SJ, Zahawi RA, Hector A (2015) Globally, functional traits are weak predictors of juvenile tree growth, and we do not know why. Journal of Ecology 103: 978-989 - doi: 10.1111/1365-2745.12401
- Salisbury CL, Potvin C (2015) Does tree species composition affect productivity in a tropical planted forest? Biotropica 47: 559-568 - doi: 10.1111/btp.12252
- Sapijanskas J, Paquette A, Potvin C, Kunert N, Loreau M (2014) Tropical tree diversity enhances light capture through crown plasticity and spatial and temporal niche differences. Ecology 95: 2479-2492 - doi: 10.1890/13-1366.1
- Schwendenmann L, Pendall E, Sanchez-Bragado R, Kunert N, Hölscher D (2014) Tree water uptake in a tropical plantation varying in tree diversity: interspecific differences, seasonal shifts and complementarity. Ecohydrology 8: 1-12 - https://doi.org/10.1002/eco.1479
- Riedel J, Dorn S, Plath M, Potvin C, Mody K (2013) Time matters: Temporally changing effects of planting schemes and insecticide treatment on native timber tree performance on former pasture. Forest Ecology and Management 297: 49-56 - doi: 10.1016/j.foreco.2013.02.003
- Sapijanskas J, Potvin C, Loreau M (2013) Beyond shading: litter production by neighbors contributes
to overyielding in tropical trees. Ecology 94: 941-952 - doi: 10.1890/12-0337.1
- Sprenger M, Oelmann Y, Weihermüller L, Wolf S, Wilcke W, Potvin C (2013) Tree species and diversity effects on soil water seepage in a tropical plantation Forest Ecology and Management 309: 76-86 - doi: 10.1016/j.foreco.2013.03.022
- Yamamura T, Schwendenmann L, Lear G (2013) Tree species identity has little impact on the structure of soil bacterial communities in a 10-year-old tropical tree plantation. Biology and Fertility of Soils 49: 819-828 - doi: 10.1007/s00374-013-0774-x
- Kunert N, Schwendenmann L, Potvin C, Hölscher D (2012) Tree diversity enhances tree transpiration in a Panamanian forest plantation. Journal of Applied Ecology 49: 135-144 - doi: 10.1111/j.1365-2664.2011.02065.x
- Plath M, Dorn S, Riedel J, Barrios H, Mody K (2012) Associational resistance and associational susceptibility: specialist herbivores show contrasting responses to tree stand diversification. Oecologia 169: 477-487 - doi: 10.1007/s00442-011-2215-6
- Plath M, Mody K, Potvin C, Dorn S (2011) Establishment of native tropical timber trees in monoculture and mixed-species plantations: Small-scale effects on tree performance and insect herbivory. Forest Ecology and Management 261: 741-750 - doi: 10.1016/j.foreco.2010.12.004
- Potvin C, Mancilla L, Buchmann N, Monteza J, Moore T, Murphy M, Oelmann Y, Scherer-Lorenzen M, Turner BL, Wilcke W, Zeugin F, Wolf S (2011) An ecosystem approach to biodiversity effects: carbon pools in a tropical tree plantation. Forest Ecology and Management 261: 1614-1624 - doi: 10.1016/j.foreco.2010.11.015
- Ruiz-Jaen MC, Potvin C (2011) Can we predict carbon stocks in tropical ecosystems from tree diversity? Comparing species and functional diversity in a plantation and a natural forest. New Phytologist 189: 978-987 - doi: 10.1111/j.1469-8137.2010.03501.x
- Schneebeli M, Wolf S, Kunert N, Eugster W, Mätzler C (2011) Relating the X-band opacity of a tropical tree canopy to sapflow, rain interception and dew formation. Remote Sensing of Environment 115(8): 2116-2125 - https://doi.org/10.1016/j.rse.2011.04.016
- Wolf S, Eugster W, Potvin C, Buchmann N (2011) Strong seasonal variations in net ecosystem CO2 exchange of a tropical pasture and afforestation in Panama. Agricultural and Forest Meteorology 151: 1139-1151 - doi: 10.1016/j.agrformet.2011.04.002
- Wolf S, Eugster W, Potvin C, Turner BL, Buchmann N (2011) Carbon sequestration potential of tropical pasture compared with afforestation in Panama. Global Change Biology 17: 2763-2780 - doi: 10.1111/j.1365-2486.2011.02460.x
- Oelmann Y, Potvin C, Mark T, Werther L, Tapernon S, Wilcke W (2010) Tree mixture effects on aboveground nutrient pools of trees in an experimental plantation in Panama. Plant and Soil 326: 199-212 - doi: 10.1007/s11104-009-9997-x
- Zeugin F (2010) The functional significance of tree diversity for nutrient acquisition in a tropical tree plantation. PhD thesis, ETH Zurich
- Zeugin F, Potvin C, Jansa J, Scherer-Lorenzen M (2010) Is tree diversity an important driver for phosphorus and nitrogen acquisition of a young tropical plantation? Forest Ecology and Management 260: 1424-1433 - doi: 10.1016/j.foreco.2010.07.020
- Potvin C, Dutilleul P (2009) Neighborhood effects and size-asymmetric competition in a tree plantation varying in diversity. Ecology 90: 321-327 - doi: 10.1890/08-0353.1
- Delagrange S, Potvin C, Messier C, Coll L (2008) Linking multiple-level tree traits with biomass accumulation in native tree species used for reforestation in Panama. Trees - Structure and Function 22: 337-349 - doi: 10.1007/s00468-007-0189-0
- Healy C, Gotelli NJ, Potvin C (2008) Partitioning the effects of biodiversity and environmental heterogeneity for productivity and mortality in a tropical tree plantation. Journal of Ecology 96: 903-913 - doi: 10.1111/j.1365-2745.2008.01419.x
- Murphy M, Balser T, Buchmann N, Hann V, Potvin C (2008) Linking tree biodiversity to belowground process in a young tropical plantation: impacts on soil CO2 flux. Forest Ecology and Management 255: 2577-2588 - doi: 10.1016/j.foreco.2008.01.034
- Potvin C, Gotelli NJ (2008) Biodiversity enhances individual performance but does not affect survivorship in tropical trees. Ecology Letters 11: 217-223 - doi: 10.1111/j.1461-0248.2007.01148.x
- Scherer-Lorenzen M, Bonilla JL, Potvin C (2007) Tree species richness affects litter production and decomposition rates in a tropical biodiversity experiment. Oikos 116: 2108-2124 - doi: 10.1111/j.2007.0030-1299.16065.x
- Potvin C, Whidden E, Moore T (2004) A case study of carbon pools under three different land-uses in Panama. Climatic Change 67: 291-307 - doi: 10.1007/s10584-004-0079-z